

A comprehensive review shows how the brain’s immune defenses combat and promote neurodegeneration and reveals why timing, cell type, and disease context are important for future treatments.
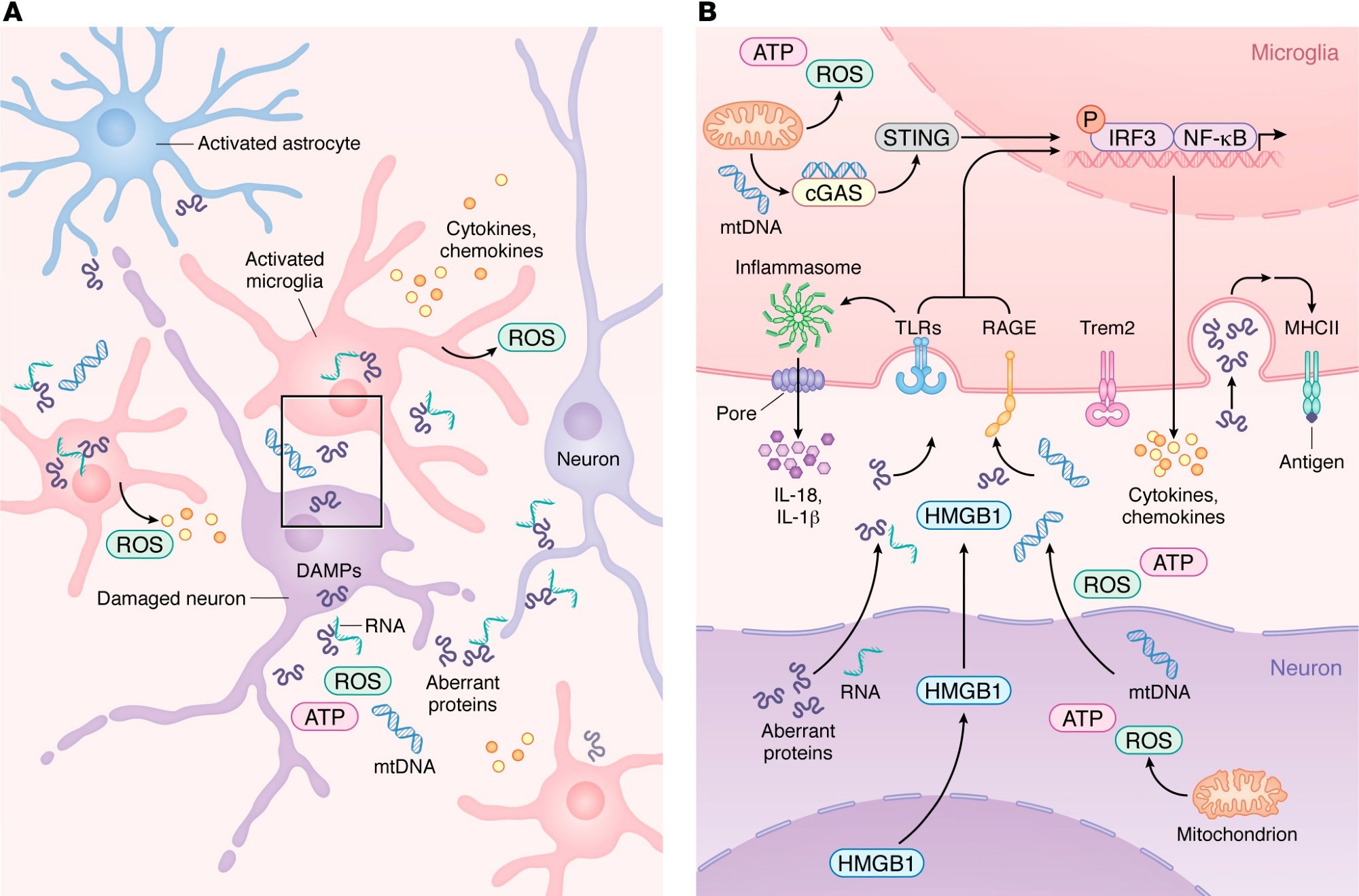
Innate immune activation and microglial migration during neurodegeneration. (a) During neurodegeneration, damaged neurons release DAMPs such as mtDNA, ROS, HMGB1, abnormal proteins, and pTau-binding RNA. These signals activate resident microglia and astrocytes, which subsequently recruit peripheral immune cells. As shown in more detail, Bactivated microglia upregulate PRRs such as TLR, RAGE, and cGAS, allowing detection of neuron-derived DAMPs. Engagement of PRRs activates the inflammasome pathway, which induces proinflammatory gene expression programs and promotes the release of chemokines and cytokines into the CNS parenchyma. As microglia transition from a homeostatic state to a reactive state in a TREM2-dependent manner, injury-associated microglia increase MHC-II expression, supporting antigen presentation and coordination of adaptive immune responses that seek to limit neurodegeneration.
Recent reviews published in clinical research journal We synthesized the current evidence regarding immune mechanisms in neurodegeneration. Accumulating evidence suggests that the immune system not only promotes the progression of neurodegenerative diseases but also contributes to their development through maladaptive cell signaling and activation. The central nervous system (CNS) parenchyma relies on resident immune cells to maintain the blood-brain barrier (BBB) under steady-state conditions and to respond to dysfunctional cells, pathogens, or cell damage.
Neurons respond to cellular dysfunction by activating signaling pathways, while microglia and astrocytes reduce danger signals and recruit peripheral immune cells. However, these responses can exacerbate aberrant protein deposition during neurodegeneration, and the balance between harmful and beneficial immune involvement is not well defined. In this review, researchers reviewed current knowledge about immune mechanisms in neurodegeneration.
Innate immune signaling in neurodegenerative diseases
Aberrant proteins such as phosphorylated tau, amyloid beta (Aβ), and alpha-synuclein have been identified as damage-associated molecular patterns (DAMPs) that activate pattern recognition receptors in the CNS. Toll-like receptor 2 (TLR2) and TLR4 are upregulated in mouse models of Alzheimer’s disease (AD) and in the brains of AD patients.
Binding of Aβ to TLR4 or TLR2 increases the expression of cytokines such as tumor necrosis factor (TNF)-α, interleukin (IL)-6, IL-1β, IL-17, and IL-10 in microglia and astrocytes. Pharmacological inhibition or genetic ablation of these TLRs exacerbates cognitive decline by reducing microglial activation and increases Aβ burden in the brain. This review also highlights inflammasomes and DNA sensing pathways, such as NLRP3 and cGAS-STING, as further contributors to neuroinflammation across neurodegenerative diseases.
Studies have consistently reported increased expression of receptor for advanced glycation end products (RAGE) on neurons, astrocytes, and microglia in AD, Parkinson’s disease (PD), and amyotrophic lateral sclerosis. RAGE overexpression in a mouse AD model accelerated cognitive impairment, whereas its deficiency decreased memory impairment.
Genetic studies of Alzheimer’s disease patients have identified two single nucleotide polymorphisms (SNPs) in trigger receptor 2 (TREM2), which is highly expressed by microglia, as risk factors comparable in magnitude to carrying the apolipoprotein E (APOE) ε4 allele. These observations indicate that microglia play an important role in the pathogenesis of neurodegeneration.
T cell responses across neurodegenerative diseases
Increasing evidence indicates that cluster of differentiation 4 (CD4+) T cells have disease-specific functions across neurodegenerative diseases. Transfusion of Aβ-restricted CD4+ T cells into a transgenic AD mouse model was neuroprotective and prevented cognitive decline. In contrast, α-synuclein-specific CD4+ T cells were neurotoxic and promoted inflammation in PD models.
Overexpression of α-synuclein increases the recruitment of CD4+ T cells that produce interferon-γ, whereas depleting these cells reduces neuronal loss. These observations indicate that CD4+ T cells may be protective in some neurodegenerative diseases and pathogenic in others. Furthermore, CD8+ T cells are receiving increasing attention as contributors to CNS pathology. CD3+ and CD8+ T cells have been detected in the hippocampus of AD, and CD3+ T cell numbers correlate with tau pathology.
Similarly, increased numbers of CD8+ T cells were observed in PD and correlated with neuronal cell death, but whether this relationship is causative or reactive remains unclear. Additionally, accumulation of neuroprotective CD8+ T cells in the brains of AD mice has been reported and shown to limit amyloid plaque growth. Overall, studies reveal that CD8+ T cells play diverse roles in neurodegenerative diseases, shaped by antigen specificity, microglial interactions, and tissue residency. This review also notes the growing interest in granzyme K-expressing CD8+ T cells as potentially specialized players in several neurodegenerative diseases.
Effects of aging, trauma, and viruses on central nervous system immunity
Aging is a major risk factor for neurodegenerative diseases. In young people, microglia survey the CNS, astrocytes maintain the BBB, and only a small number of T cells enter the parenchyma. However, as we age, chronic low-grade inflammation occurs, during which many immune pathways become dysregulated.
Additionally, repeated head injuries can accelerate neurodegeneration. Traumatic brain injury, even when occurring during childhood, is associated with the development of PD, AD, and chronic traumatic encephalopathy. After mild traumatic brain injury, the release of DAMPs by injured cells triggers microglial activation, which clears debris and strengthens weakened BBB areas.
Repeated injury can increase reactivity and cause microglial death, thereby allowing myelomonocytic cell invasion. Similar to repetitive injuries, viral infections can alter central nervous system immunity and influence vulnerability to neurodegeneration. Recent studies have associated non-neurotropic and neurotropic viral infections with a higher risk of neurodegenerative diseases.
Immune timing and its impact on neurodegeneration
Taken together, neurodegenerative diseases result from a complex interplay of environmental and biological factors that govern the magnitude and timing of immune activation and the programming of T cells, microglia, and other leukocytes. Viral infections, repetitive head trauma, and aging remodel the CNS immune environment. Many of the immune pathways involved exhibit stage-dependent and context-specific functions.
Microglial activation initially helps remove abnormal proteins, but can become maladaptive if stimulation is prolonged. Similarly, CD8+ T cells can promote neural injury or support tissue repair based on antigen specificity, local signaling cues, and effector programming. Therefore, understanding the cell-intrinsic state and temporal dynamics of immune cells is important for understanding immune signatures and determining when immunomodulatory interventions are beneficial.